Zur Statik und zum Gleichgewicht von Theropoden
und die Widerlegung der Metatarsus-Theorie von Glen Jay Kuban
Von Dipl.-Ing. Hans-Joachim Zillmer
Zweibeinige Dinosaurier liefen auf Ihren Zehen, was den in den meisten Fällen einen abgerundeten Hinterteil eines Abdrucks verursachte. Insbesondere im Bereich des Paluxy River bei Glen Rose in Rose in Texas wurden jedoch einige Trittsiegel entdeckt, die eine nach rückwärts verlängerte Form von bis zu über 50 Zentimeter Fußlänge aufweisen und an eine menschliche Fußform erinnern. Diese Erscheinungsform hat in der Vergangenheit zu kontroversen Diskussionen geführt. Eine 1986 von Glen Jay Kuban (1) vorgestellte Hypothese soll plausibel machen, dass die längliche Form von zweibeinigen Dinosaurier stammen, die anstatt auf dem Vorderfuß ihren Körper teilweise unmittelbar auf dem Mittelfuß (Metatarsus) gingen. Die Abstützung des Körpergewichts auf dem Metatarsus (Mittelfuß) anstatt auf dem mit drei Krallen bewehrten Vorderfuß ist aus anatomischen und statischen Gründen jedoch nicht möglich.
Abb. 1. Rekonstruktion von Theropoden früher (links) und heute Tyyrannosaurus rex im Museum of Natural History in New York).
Über einhundert Jahre hinweg wurden zweibeinige Dinosaurier (Theropoden) mit hoch erhobenem Kopf auf zwei Beinen stehend zur Schau gestellt (Abb. 1 links). Dabei lag der Schwanz bei den in den Museen aufgebauten Skeletten auf dem Boden. Auf diese Art wurde der Stand der Dinosaurier stabilisiert und bildete statisch gesehen ein dreidimensionales Dreibein. Da Pfade mit Trittsiegeln von Dinosauriern nie Schwanzschleifspuren zeigen, werden aktuelle Rekonstruktionen z. B. von Fleischfressern wie Allosaurus oder Tyrannosaurus rex und Pflanzenfressern wie Iguanodon mit im Prinzip waagerecht gehaltener Wirbelsäule neu aufgebaut.
In Deutschland wurden beim Pflanzen fressenden Plateosaurus die Schwanzknochen neu zusammengesetzt, sodass dieser Saurier seiner möglichen Fortbewegungsart als Zweibeiner bei auf dem Boden schleifenden Schwanz beraubt wurde und neuerdings als Vierbeiner mit hochgehaltenem Schwanz rekonstruiert wird. Der Plateosaurus im Paläontologischen Museum München wurde 2002 in ganz neuer Skelettkonstruktion mit waagerecht gehaltener Wirbelsäule wieder errichtet ( siehe auch ).
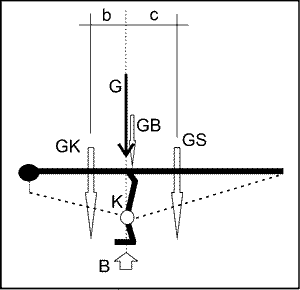
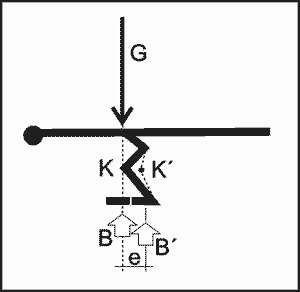
Im Gegensatz zu Plateosaurus, der durch den vierbeinigen Stand größere Standfestigkeit erlangte, haben auf zwei Beinen laufende Theropoden ein statisches Problem, denn der Körper einschließlich des Kopfes sollte sich in der Standphase mit dem Schwanz im Gleichgewicht befinden, also wie bei einem Kran austariert sein. Dieses Gleichgewicht ist hergestellt, wenn die Momente, gebildet aus Kraft mal Hebelarm, links und rechts vom Drehpunkt, also dem Punkt der Unterstützung, gleich sind. Dieser Punkt sollte auf einem senkrechten Vektor bzw. auf einer Achse liegen, in der auch der Schwerpunkt des Tiergewichts (G) sowie der durch das Gesamtgewicht des Tieres erzeugte Bodendruck als resultierende Stützungs- bzw. Reaktionskraftgröße (B) wirkt. Auf dieser Achse, die wir Ruheachse nennen wollen, liegt im Ruhestand auch das Kniegelenk (K), um (kraftaufwendige) Verdrehungen im Zustand des Gleichgewichts zu vermeiden. Bei Einhaltung dieser Bedingungen ist die geringste Muskel-, Sehnen- und Bänderbean-spruchung gegeben.
Da jetzt die Hebelarme für das Gewicht des Körpers (GK) und des Schwanzes (GS) bekannt sind, erhalten wir eine Gleichgewichtsbeziehung, falls der Schwerpunkt des Beingewichts idealisiert in der Ruheachse liegt und kein Drehmoment (M) erzeugt, da der Hebelarm gleich Null ist:
GK x b = GS x c oder M = 0 = GS x c – GK x b
Abb. 2. Idealisierter Theropode im Gleichgewichts-Ruhestand. G = Gesamtgewicht, GK = Gewicht Körper, GS = Gewicht Schwanz, GB = Gewicht Beine, K = Kniegelenk, B = Unterstützungskraftgröße im Boden.
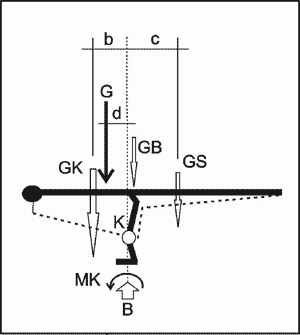
Die modernen Rekonstruktionen zeigen jedoch ein anderes Bild, denn der Schwanz wird sehr schlank dargestellt und kann kein Gegengewicht zum schweren Körper bilden. Die gestrichelten Linien zeigen die ungefähre Gewichtsverteilung, die sich aufgrund heutiger Rekonstruktionen ergibt (Abb. 3). Entsprechend wirkt das Gesamtgewicht aus GK und GS nicht in der Ruheachse, sondern erzeugt ein Kippmoment (Gewicht der Beine GB vernachlässigt),
MK = G x d = GK x b – GS x c
das im Kniegelenk K wirken würde und ausgeglichen, also stabilisiert werden müsste. Das Kippmoment MK wirkt aber auch auf die Fußflächen bzw. müsste über die Füße in die Aufstandsfläche bzw. den Untergrund abgetragen werden, da der Theropode sonst (ohne Aus-gleichsreaktion) auf die Nase kippt. Die Wirkungslinie (= Resultierende) des Gewichts G verläuft außerhalb der Fußflächen (= Aufstandsfläche). Deshalb ist für den Ruhestand kein Gleichgewicht gegeben. Der Theropode wäre gezwungen, einen Ausfallschritt zu machen, um überhaupt die Chance zu erhalten, den Stand zu stabilisieren. Dies würde, ob mit oder ohne Bewegung, eine permanent aktive Muskelarbeit bedeuten. Ein Ruhestand mit nebeneinander gestellten Beinen, also ohne größere Muskelarbeit, ist nach den aktuellen Skelettrekonstruktionen nicht möglich.
Abb. 4. Idealisierter Theropode gemäß aktueller Rekonstruktion mit massigem Körper und schmalem Schwanz: Ein Ruhestand in Form eines Gleichgewichts ist nicht möglich
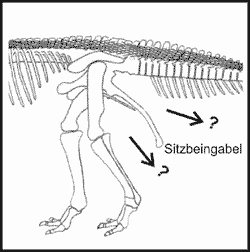
Abb. 3. Lag die Afterknolle (?) unter oder über der Sitzbeingabel? Das Rückgrat ist durch Sehnen versteift(vgl. Abb. 5).
Um dem Theropoden eine gewichtsmäßig ausgeglichene Ruhestellung zu geben, sollte die Gewichtsverteilung – wie in der Abb. 2 abstrahiert skizziert (gestrichelte Linie) – angeordnet sein. Dies bedeutet einen wesentlich höheren und damit schwereren Schwanz. Die aus statischer Sichtweise falsche Rekonstruktion der Theropoden ergibt sich aus der Platzierung der Afterknolle. Diese Körperöffnung wird allgemein über der Sitzbeingabel vorgesehen, wodurch für den Schwanz nur noch eine geringe Höhe zur Verfügung steht (Abb. 3).
Da man bis auf wenige Ausnahmen nur Skelett- aber kaum Weichteilreste und keine komplett versteinerten Tiere findet, ist die Platzierung der Afterknolle Spekulation. Diese kann sich alternativ unten in der Nähe des tiefsten Körperpunktes befunden haben (2).
Dadurch kann insgesamt eine vollere Körperform mit einem hohen Schwanz-ansatz und damit einem höheren sowie schwereren Schwanz entstehen (3).
Auf einen wesentlich höheren als bisher rekonstruierten Schwanz deuten die selten zu findenden und bei Rekonstruktionen oft nicht berücksichtigten Knochenanhänge (Chevrons) hin, die sich unterhalb des Schwanzes befinden, um so den Schwanz als eine versteifte Scheibe auszubilden. Die Folge ist, dass der Schwanz sich nicht in vertikaler Hinsicht durchbiegen kann, und damit nur in horizontaler beweglich ist.
Diese Steifigkeit des Schwanzes ist erforderlich, um die Übertragung des Schwanzgewichtes hin zu den Beinen als Kontergewicht zum Körpergewicht zu gewährleisten und andererseits der Beobachtung genüge zu tun, dass keine Schwanzschleifspuren während des Gehvorganges entdeckt werden.
Die Rekonstruktion von Theropoden als auszutarierende, wippende Konstruktion erscheint allein aus statischen Gesichtspunkten zweifelhaft, insbesondere da die Rekonstruktion des Körpers dieser Tiere immer voluminöser ausfällt und damit trotz schwererem Schwanz kein Gleichgewicht gewährleistet ist.
Abb. 5. Knöcherne Sehnen verstärken statisch den Schwanz (als Zuggurt) und versteifen diesen gegen vertikale Durchbiegungen. Die Chevrons hängen unter dem Rückrat, wie bei diesem Corythosaurus, und bewirken einen hohen Schwanzquerschnitt. Oval = Afterknolle
Abb. 6. Ohne Bauchrippen rekonstruierte Theropoden mit zu hoch aufgesetzten Schultergürtel und kurzen Armen.
Die meisten Dinosaurier werden außerdem nur mit Rippen, aber nicht mit den erforderlichen Bauchrippen rekonstruiert, die äußerst selten fossil erhalten blieben. Ohne die Anbringung von Bauchrippen erscheint der Körper von Theropoden schlanker und die Vorderfüße werden über den Schultergürtel (Scapula und Coracoid) oben auf die Rippen in der Nähe der Wirbelsäule befestigt (Abb. 6). Dadurch bekommt beispielsweise Tyrannosaurus rex besonders kurze, funktionslos erscheinende Arme.
Im Natural History Museum Washington D. C. wurde der angebliche Raubsaurier Allosaurus fragilis, ein früher Verwandter der Tyrannosaurier, komplett mit Bauchrippen rekonstruiert, wodurch ein plumperes Aussehen entsteht (Abb. 7). In diesem Fall wurde der Schultergürtel nicht oben auf den Rippen, sondern unten an den Bauchrippen befestigt. Erst dadurch entsteht ein kraftschlüssiger Verbund der Vorderarme mit dem Skelett, der bei Anordnung der Arme gemäß Abb. 6 fehlt.
Außerdem hängen bei der Berücksichtigung von Bauchrippen nach Abb. 7 die kurzen Vorderarme weit unter dem Bauch und reichen bei horizontaler Haltung der Wirbelsäule fast bis zum Boden hinab. Damit entsteht fast ein Vierfüßer, der sich zumindest bei Bedarf mit den Vorderbeinen abstützen konnten, um das nach vorne Kippen zu verhindern bzw. das Kippmoment (MK) auszugleichen.
Oder waren Theropoden auch Vierfüßer, genauso wie Iguanodon und Corythosaurus, die bis vor kurzem noch als Zweibeiner rekonstruiert wurden ( 4 ) ?
Abb. 7. Allosaurus mit Bauchrippen und tief herabhängenden, fast auf dem Boden schleifenden Armen.
Abb. 8. Bein von Tyrannosaurus rex
Falls Theropoden jedoch Zweifüßer sein sollen, können Sie aufgrund ihres großen Körpergewichts keine flinken Räuber gewesen sein, da diese Saurier allein aus statischen, aber auch dynamischen Gründen mit dem Ausbalancieren des Gleichgewichts beschäftigt sein mussten.
Zweibeinige Saurier liefen auf ihren Zehen, wie versteinerte Spuren zeigen. Insbesondere am und im Paluxy River bei Glen Rose in Texas wurden einzelne Trittsiegel mit verlängerter Form entdeckt, die wie menschenähnliche Fußabdrücke aussehen, mit einer Gesamtlänge von über 50 Zentimetern. Glen Jay Kuban erklärte, dass zweibeinige Dinosaurier teilweise auf dem aus Röhrenknochen bestehenden Mittelfuß (Metatarsus) gingen1. Ohne weitere Überprüfung oder Untersuchung diente diese Erklärung als Beweis gegen alle seit 1908 und auch zukünftig zu entdeckenden »Menschenfußspuren« (5).
Kann ein Theropode auf seinem Metatarsus, also gleichzeitig auf dem Vorder- und Mittelfuß (Abb. 8) gehen, um derart die verlängerten Trittsiegel zu erzeugen? Analog der bisherigen statischen Betrachtung sollte das Knie in der Ruheachse liegen (Abb. 2), auch in dem Augenblick der vollen Gewichtsverlagerung von einem Fuß auf den anderen, um die Beanspruchung von Muskeln, Bändern und Knochen auf ein Minimum zu reduzieren.
Bei einem Zweibeiner muss das Gewicht vom Kniegelenk aus durch Unterschenkel und Fuß in den Boden geleitet werden. Die Abweichungen der Gelenke von der Ruheachse sollten so klein wie möglich sein (Abb. 2 vgl. Abb. 8). Auch das Gelenk zwischen Vorder- und Mittelfuß sollte in der Nähe der Ruheachse liegen. Wird diese Bedingung eingehalten, werden zusätzliche Drehmomente (Verdrehungen) auch in diesem Gelenk vermieden bzw. auf ein Minimum beschränkt.
Bei normaler Fußhaltung bildet der Mittelfuß (Metatarsus) zur Bodenfläche einen Winkel von ungefähr 70 Grad. Würde der Theropode den Mittelfuß horizontal auf den Boden pressen, dann entsteht, falls das Knie in der ursprünglichen Ruheachse verbleibt, ein Winkel von ungefähr 135 Grad (vgl. Abb. 10). Eine Überdehnung der Bänder und ein Bruch des Gelenks zwischen Mittelfuß und Unterschenkel wäre die Folge.
Die dreizehigen Trittsiegel am Paluxy River sollen von Acrocanthosaurus stammen. Da die langen Trittsiegel eine Länge von 55–65 cm und die normalen Zehenabdrücke des Vorderfußes von 35–40 cm aufweisen, ergibt sich ein möglicher minimaler Hebelarm, falls der resultierende Bodendruck B in Mitte Vorderfuß wirkt, von k = (55–35) + 35/2 = 37,5 cm (Abb. 9).
Acrocanthosaurus soll zwei bis drei Tonnen gewogen haben. Es ergibt sich ein zusätzliches Moment aus Gesamtgewicht mal Hebelarm, falls das Tier im Gleichgewicht stehen soll, von mindestens
MG = G x k > 20.000 x 0,375 = 7.500 Nm
Dieses wirkt zusätzlich zum Gesamtgewicht im Gelenk zwischen Mittelfuß und Unterschenkel und müsste über den Mittelfuß übertragen werden. Umgerechnet bzw. aufgelöst in ein Kräftepaar von Druck- (D) und Zugkraft (Z) bei einem geschätzten wirksamen Hebelarm im Gelenk von – da Bänder nicht fossil erhalten – von 0,2 m:
Z = –D = M / n = 7.500 / 0,2 = 37.500 N.
Schätzen wir den Querschnitt des nicht fossil erhaltenen Muskels bzw. des Bandes auf 50 cm2, dann ergibt sich eine Spannung S aus Kraft geteilt durch Fläche von
S = F / A = 37.500 N / (50 x 10^-4) m^2 = 7.500.000 N/m^2 = 75 bar
Abb. 9. Auftritt mit Vorderfuß (F) und Metatarsus (M). Bodenkraft: im Ruhestand (B), beim Metatarsus-Tritt (B´).
Diese Spannung addiert sich zu der Vorspannung des Muskels, die durch die Überdrehung des Gelenks auf 135 bis 140 Grad (Abb. 10) entsteht. Die Übertragung von Zug- und Druckkräften dieser Größenordnung, zusätzlich zu dem auch durch das Gelenk zu übertragenden Gesamtgewicht des Tieres G, ist undenkbar. Gelenke können naturgemäß keine bzw. nur relativ geringe Momente (Kraft x Hebelarm) über die Muskeln, Sehnen und Bänder übertragen. Im Falle des Auftritts auf dem Mittelfuß M (Metatarsus) müßte im Gegensatz zum Ruhestand auf dem Vorderfuß F (Abb. 9) fast das dreifache (oder mehr) der normalen Belastung übertragen werden. Bewegt sich der Theropode, kommen weitere enorme Trägheitskräfte hinzu.
Außerdem ist das Gelenk zwischen Mittelfuß und Unterschenkel und auch der aus Röhrenknochen bestehende Mittelfuß (Metatarsus) konstruktiv nicht zur Übertragung von hohen Kräften auf den Boden geeignet. Röhrenknochen sind zwar in Längsrichtung sehr tragfähig, ähnlich wie Eichenholz, bei Beanspruchung in Querrichtung, wie es beim Auftritt auf den Mittelfuß (Metatarsus) der Fall wäre, ist die Tragfähigkeit wesentlich geringer anzusetzen, da der Knochen auf Biegung beansprucht wird.
Abb. 10. Haltung des Theropoden beim Auftritt mit Vorderfuß und Mittelfuß.
Ein Gleichgewicht ist beim Tritt auf dem Mittelfuß nicht möglich, unabhängig von der Kniestellung. Dieses könnte nur über eine Ausgleichsbewegung erzielt werden, wobei der Zeitpunkt des Anhebens eines Beines bei der Vorwärtsbewegung sowie der erforderlichen Gewichtsverlagerung nach vorn für das Tier gefährlich wäre, da ein weiteres zusätzliches Kippmoment entsteht
Es ist auch zu beachten, dass die getrennte Anordnung von Vorder- und Mittelfuß den Sinn hat, neben der Beweglichkeit über die Muskeln und Bänder eine Dämpfung des Bewegungsapparates zu gewährleisten, damit ruckartige Belastungen des Skeletts, wie durch abgefederte Autoräder, vermieden werden.
Tritt der Theropode direkt auf seinen Mittelfuß, wird der Aufprallschock beim Auftritt ohne Dämpfung auf das ganze Tier übertragen. Freiwillig würde der Theropode deshalb nicht auf seinen Mittelfuß treten. Außerdem ist ein sehr großer Energieaufwand nötig, um den Mittelfuß (Metatarsus) aus rückwärtiger (horizontaler) Position gegen das Eigengewicht um ungefähr 70 Grad (siehe Abb. 8) wieder aufzurichten und in Normalstellung (Ruhestand) zu bringen.
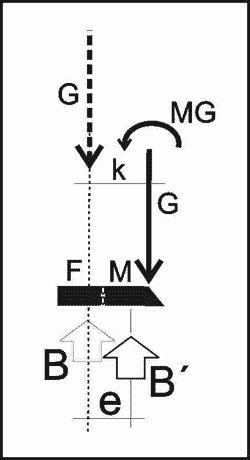
Betrachten wir jetzt alternativ den anderen Fall, dass, um eine Überdrehung und Überbelastung des Gelenks zwischen Mittelfuß und Unterschenkel zu vermeiden, es denkbar wäre, das Knie während des Metatarsus-Auftritts nicht in die Ruheachse zu bringen, sondern in einem steileren Winkel von 100 Grad anstatt 135 Grad zu belassen. Das Knie nimmt dann die Position K´ anstatt K ein (Abb. 10). Befindet sich das Kniegelenk beispielsweise in der Achse der Unterstützung B´, würden wiederum zusätzliche Momente (Eigengewicht mal Hebelarm) entstehen, die nicht übertragen werden können (Abb. 10):
M = G x e > zulässig M
Bei der Fortbewegung kommen zur Kraftverteilung im Ruhestand weitere Trägheitskräfte hinzu, durch die zusätzlich auch horizontal wirkende Kräfte entstehen und wirken.
Unabhängig von der zusätzlichen Belastung der Knochen und Muskeln würde durch den Auftritt auf dem Mittelfuß ein größeres Kippmoment als beim Auftritt auf den Vorderfuß entstehen, da der Schwerpunkt der Bodenbelastung mit dem Hebelarm e (Abb. 10) weiter vom Schwerpunkt des Tiergewichts G entfernt ist. Das Kippmoment kann nicht über die Aufstandsfläche der Füße übertragen werden, da der Schwerpunkt der Belastung weit außerhalb, also sehr weit vor den Fußflächen (= der das Tiergewicht im Boden stützenden Fläche) liegt.
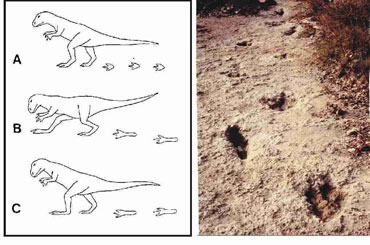
Abb. 11. Haltung und Gang eines Theropoden nach Kuban (1):
A = Gang auf Vorderfuß, B und C unnatürlicher Gang auf dem Mittelfuß, der großen Energieaufwand und große Verdrehungen der Gelenke bewirkt. Das rechte Bild zeigt einen Dinosaurier-Pfad mit einem länglichen Trittsiegel. Aus Kuban (1).
Untersucht man die langen Trittsiegel, die angeblich durch den Tritt auf den Mittelfuß entstanden sein sollen, dann fällt auf, dass diese nur sehr vereinzelt innerhalb eines aus einzelnen Fußspuren bestehenden Pfades zu finden sind. Der Verfasser hat viele Fossilienstätten besucht und nirgendwo, außer am Paluxy River, derart längliche Trittsiegel mit einer Gesamtlänge von über 50 cm gesehen. Auch wenn es diese bei anderen Fundstätten geben sollte, handelt es sich um sehr seltene Funde.
Nach Kuban1 weist der in Abb. 12 (links) dokumentierte Abdruck, oberflächlich betrachtet, eine menschliche Form auf. Er soll aber ein Metatarsus-Trittsiegel sein. Eine nähere Erklärung wird nicht angeboten. Die schmale Form soll entstanden sein, als der ursprünglich weiche Matsch nach der Verdrängung wieder zurückfloss.
Vom Abdruck der drei Zehen ist nichts zu sehen. Kuban (1) behauptet: »Auf einem schlammigen Boden könnte auch ein Zurückfließen des Schlamms über den vorderen Teil einer Spur die Zehenabdrücke wieder ausfüllen und zudecken.
Nach der Erhärtung muss das obere Sediment zuerst erodieren, bevor die Zehenadrücke sichtbar werden (6)«. Warum passierte dies nur bei dem zweiten Abdruck in der Abb. 11, aber nicht bei den anderen? Warum wurden die Krallen gerade bei diesem Abdruck komplett und bei den anderen überhaupt nicht abgedeckt? Falls der den Abdruck umgebende Schlamm in diesen hinein fließen würde, müsste dies an der Form der Oberfläche des Bodens zu sehen sein,
Abb. 12. Der längliche Abdruck (links) soll nach Kuban (1) ein Metatarsus-Trittsiegel ohne sichtbaren Vorderfuß sein, der angeblich durch den Schlamm zusammengedrückt wurde. Von einem solchen Vorgang ist im Bodenrelief nichts zu erkennen. In der rechten und mittleren Abbildung ist deutlich die zerstörte Bodenstruktur durch Nachfließen des Schlamms zu erkennen, wobei im mittleren Bild der ursprünglich verdrängte Schlamm die Abdrücke der Krallen fast komplett wieder schloss, aber ein entsprechendes Muster hinterließ. Originalfotos: Kuban. Falschfarbenbearbeitung: Zillmer
denn es sollte eine kleine Senke oder ein Riss in der Bodenfläche entstehen. Da dies nicht beschrieben wird, kann man davon ausgehen, dass – wie auf der Abbildung 11 (rechts) und 12 (links) erkennbar – sich keine Formänderung der den Abdruck umgebenen Oberfläche ereignet hat. Durch optische Verfahren wie die Anwendung einer Temperaturkarte können Unterschiede deutlicher gemacht werden. Der Abdruck in Abb. 12 (links) zeigt eine homogene Struktur des umgebenen Felsens, während der Abdruck in Abb. 12 (Mitte) eine gebrochene Bodenstruktur zeigt und zwar in dem Bereich, wo der Schlamm in den hinteren Teil des Abdrucks tatsächlich floss. Entsprechendes ist bei dem rechten Bild der Abb. 12 zu erkennen, wo sich der oberhalb links des Abdruck befindliche Teil, der eigentlich die Krallen zeigen müsste, ablöste und auf diese Art der entstandene Riss den Abdruck einer einzelnen Kralle vorgespiegelt.
Abb. 13. Dieses lange Trittsiegel (Foto 1) gilt als ein Paradebeispiel für ein Metatarsus-Trittsiegel (Foto 1 und Negativfoto 2). Es handelt sich hier jedoch um zwei verschiedene Abdrücke (Foto 1oben bzw. 2 oben und Foto 5 bzw. 4), die zufällig hintereinander liegen, siehe zusammengesetztes Foto 3 (= Foto 1 oben) sowie Foto 4,das ein Negativbild von Foto 5 darstellt. Das Originalfoto 2 unten als Negativbild und Foto 4 (= Foto 3 unten bzw. Foto 5) weisen die gleiche Struktur auf. Das angeblich aus einem Abdruck bestehende Metatarsus-Trittsiegel besteht aus zwei verschiedenen Trittsiegeln, die in Foto 6 sogar ineinander liegen (vgl. Pfeile in Foto 6, 4 und 2 unten). Foto 1, 5 und 6 aus Kuban, Ergänzungen: Zillmer.
Von Krallenabdrücken ist weder im linken noch im rechten Bild der Abb. 12 etwas zu erkennen. Handelt es sich beim linken und rechten Bild der Abb. 12 um Metatarsus-Trittsiegel ohne den Vorderfuß? Können Theropoden einerseits den Mittelfuß und andererseits dann auch noch den Vorderfuß des Beines derart in unnatürliche Pose abknicken, das er keine Spur hinterlässt?
Handelt es sich bei den rechten und linken Bildern der Abb. 12 um andersartige, isolierte Abdrücke? Betrachtet man ein angebliches Paradebeispiel für ein zusammen mit dem Abdruck des Vorderfußes erzeugten Metatarsus-Trittsiegel (Abb. 13/1), dann scheint dieser lange Abdruck aus zwei kleineren zu bestehen: vorne ein normaler mit drei Zehen und dahinter ein länglicher Abdruck, der aus zwei Vertiefungen mit einem dazwischen liegenden höheren Steg besteht. Die Abb. 13/2 ist eine invertierte Vergrößerung des linken Fotos. Einzelheiten sind hier deutlicher zu erkennen.
Der Abdruck aus der Abb. 12 (links) wurde vom Autor ausgeschnitten, zur Verdeutlichung der Strukturen elektronisch mit Falschfarbenbearbeitung aufbereitet, und zum Vergleich neben das angebliche Metatarsus-Trittsiegel, dem hinteren Teil des langen Abdrucks in Abb. 13/1, gesetzt. Deutlich zu erkennen ist, dass es sich hier um die gleiche Grundstruktur handelt, wie diese zuvor beschrieben wurde. Die Abb. 13/3+4 ist eine Kombination aus dem Abdruck des Vorderfußes aus Abb. 13/1, kombiniert mit dem Abdruck aus Abb. 12 (links) bzw. Abb. 13/5.
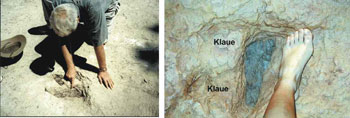
Abb. 14. Der Autor besichtigt einen länglichen Abdruck neben einem dreizehigen Trittsiegel in Barkhausen bei Bad Essen (Niedersachsen).
Es zeigt sich, dass sich hier im Vergleich mit der Abb. 13/2 (= invertierte Abb. 13/1) wiederum die gleichen Grundstrukturen zeigen. Die angeblichen langen Trittsiegel, die vom Auftreten der Theropoden auf Ihren Mittel- und Vorderfuß stammen sollen, entpuppen sich als zwei isolierte Abdrücke, die in Abb. 12 als isolierte Abdrücke dokumentiert wurden. In Abb. 13/1 liegen diese zwei Abdrücke zufällig hintereinander, während sie in Abb. 13/6 sogar ineinander liegen: Der vordere Teil des angeblichen Metatarsus-Trittsiegel liegt inmitten des dreizehigen Abdrucks (oberer Pfeil in Abb. 13/6).
Es zeigt sich, dass einige Wissenschaftler seit 1986 die Metatarsus-Erklärung der langen Trittsiegel am Paluxy River dankend als »natürliche« Erklärung für die verblüffende Ähnlichkeit dieser Abdrücke mit menschlichen Fußspuren angenommen haben und zwar ohne jedwede weitere Prüfung, schon gar nicht vor Ort! Im Internetlexikon Wikipedia wird entsprechend behauptet, dass die Wissenschaft mit den menschenähnlichen Trittsiegeln aus der Zeit der Dinosaurier kein Problem habe.
Abb. 15. Im Jahr 2001 wurden unter einer Kalksteinschicht drei neue Fußspuren von Menschen entdeckt, hier um 90 Grad gedreht gegenüber der dem Dinosaurier-Pfad. Das rechte Bild zeigt eine Vergrößerung mit dem Fuß der Tochter des Autors im Vergleich (9)
Es wurde jedoch gezeigt, dass Theropoden wie Arcocanthosaurus, Allosaurus oder Tyrannosaurus rex allein aus statischen, aber auch anatomischen Gründen nicht auf dem Mittelfuß (Metatarsus) gehen konnten. Entsprechend entpuppten sich die langen Trittsiegel als aus zwei verschiedenen Abdrücken bestehend.
Menschliche Fußabdrücke wurden in Schichten aus dem Erdmittelalter – und sogar älteren Zeitepochen – nicht nur seit 1908 am Paluxy River, sondern auch in mehreren US-Bundesstaaten zu verschiedenen Zeitpunkten dokumentiert (7).
Dabei wurden vereinzelt nicht nur Abdrücke in rudimentärer menschlicher Fußform, sondern auch mit dem Abdruck von Zehen dokumentiert (8).
Der Autor behauptet nicht, dass z. B. alle für Dinosaurier untypische längliche Spuren (vgl. Abb. 14) von Menschen stammen. Zusätzlich zu menschlichen Fußspuren werden unzweifelhaft Abdrücke in länglicher und manchmal mehr runder Form neben dreizehigen Trittsiegeln gefunden, deren Herkunft wissenschaftlich nicht geklärt ist. Der Autor schlägt als Lösung für dieses Problem vor, aus statischen Gründen ein Abstützen der Theropoden durch die Vorderbeine zu berücksichtigen (vgl. Abb. 7), damit der Körper während der Bewegung bei Bedarf im Gleichgewicht gehalten werden konnte. Dies geschah durch die nach innen gekrümmten Vorderhände (= Füße?). Die isolierten Abdrücke stellen damit Abdrücke der Handrücken dar, die manchmal in, hinter oder neben den Abdrücken der Hinterbeine platziert sind.
Abb. 16. Nebeneinander versteinerte Abdrücke der Hände und Füße von Theropoden im Seegrund bei Tuba City. H = Handabdruck (links offene Hand, rechts Handrücken). F = versteinerter Fußabdruck.
In manchen Fällen wurden entsprechende Handabdrücke auch mit geöffneter Hand erzeugt, wodurch der Eindruck entsteht, dass ein Jungtier mit der Mutter läuft, falls man davon ausgeht, dass Theropoden nur Abdrücke Ihrer Füße hinterließen.
Trotz dieser »statischen« Hilfe, des Abstützens durch die Arme, erscheint es fraglich, ob die großen (angeblich) zweibeinigen Fleischfresser-Dinosaurier überhaupt das Gleichgewicht halten konnten. Waren die großen Dinosaurier eventuell Wasser- bzw. Sumpfbewohner, wie es früher auch die Meinung der Wissenschaft war?
Denn bei Ausgrabungen in Colorado hat der Autor neben Resten von Dinosauriern in einer Art Massengrab auch Schildkröten und Krokodile ausgraben können. Im Westen von Amerika findet man diese Gemengelage in vielen fossilen (angeblichen kleinen) »Wasserlöchern«, wie z. B. im Cleveland-Lloyd Dinosaur Quarry nahe Price in Utah.
Auch die deutsche Expedition 1909 fand in Tansania (Ostafrika) Überreste von Barosaurus (vormals: Gigantosaurus) zusammen mit Muscheln, Schnecken, Belemniten und Fischen am Tendaguru, in Ablagerungen eines küstennahen Meeres der Kreidezeit. Insgesamt liegen sogar drei vom Meer abgelagerte Schichten von je 20 bis 30 Zentimetern Stärke mit Saurierresten übereinander (»Deutsches Kolonial-Lexikon«, 1920, Band III, S. 475 f.; vgl. Fraas, 1909).
Abb. 17. Fortlaufend hintereinander liegende Abdrücke von Füßen und Handrücken (H) bei Morrison in Colorado.
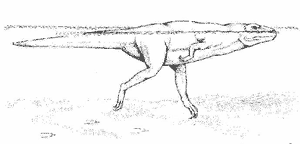
Die Art mehrerer 1980 gefundener Abdrücke verrät, dass der Fleischfresser Megalosaurus schwimmen können musste, da nur Zehen und Krallen deutlich zu erkennen sind, nicht aber der Fuß selbst. Damit konnte die Ansicht revidiert werden, dass sich Pflanzen fressende Dinosaurier (Sauropoden) einer angeblichen Bedrohung durch Flucht ins Wasser entziehen konnten (Mossmann/Sarjeant 199710).
Lebten Dinosaurier hauptsächlich in Sümpfen und Seen – auch oder gerade wegen ihrer ungeheuren Größe? Für diesen Fall hatten diese Tiere auch kein Problem mit dem Gleichgewicht. Berücksichtigt man entgegen der Darstellung in Abb. 18 jedoch analog Abb. 7 längere, fast bis auf den Boden herunter hängende Vorderarme, dann käme ein Schwimmstil vielleicht wie bei Hunden infrage. Damit könnte erklärt werden, warum Abdrücke der Hinter- und Vorderbeine isoliert, in- oder nebeneinander gefunden werden.
Der Autor konnte aufgrund der anhaltenden Erosion den alten in Abb. 11 gezeigten Dinosaurier-Pfad in Glen Rose nicht mehr vor Ort untersuchen. Jedoch scheint der zweite Abdruck dieser Serie von dem Arm (=Vorderbein?) zu stammen, während der Abdruck des Hinterbeins an dieser Stelle anscheinend fehlt. Bewegte sich das Tier schwimmend im Wasser ist dieses Phänomen erklärbar, während ansonsten ein ungeklärtes Problem entsteht, falls dieser Acrocanthosaurus sich an Land bewegt haben soll.
An Land hätten diese Tiere auch Schwierigkeiten gehabt, sich bei dem schlammigen, glitschigen Untergrund im Gleichgewicht zu halten. Es gibt nur sehr selten Spuren, die von einem Rutschen des Fußes zeugen. Sind die vielen erhalten gebliebenen versteinerten Spuren wirklich an Land in Schlammschichten erhalten geblieben?
Im Wasser ist naturgemäß eher eine Schlammschicht vorhanden und das Wasser schützt diese Spuren auch noch gegen Erosionseinflüsse, denn auf dem Trockenen erzeugte Spuren verwischen (erodieren) naturgemäß sehr schnell, auf jeden Fall schneller als die Versteinerung eintritt! Schließlich kann man vielleicht die besten Dinosaurier-Trittsiegel der Welt noch heutzutage unter Wasser im Flussbett des Paluxy River bewundern.
Abb. 18. Schwimmender Megalosaurus (10)
Tatsächlich hat man auch an der Decke von Kohleflözen Dinosaurier-Trittsiegel in verschiedenen Minen in großer Zahl gefunden. Diese Tiere sanken tief in die damals noch weiche Kohle ein und man stellt sich vor, dass die weiche Kohle sich am Boden von Sümpfen befand.
Diese offizielle Erklärung erscheint plausibel. Stellt man sich dieses Szenario ohne weiche Kohle, dafür aber mit einer schlammigen Kalkschicht am Boden des Gewässers vor, dann könnten mehrere Rätsel gelöst sein: Die Theropoden hatten keine Gleichgewichtsprobleme, da die Bewegung im Wasser statisch gestützt wird. Etliche menschlich aussehende Trittsiegel von Dinosauriern stammen tatsächlich von den Händen (=Vorderfüßen) dieser Urzeitgiganten.
Abb. 19. Dreizehige Trittsiegel an der Decke eines Kohleflözes in der Mine Castle Gate in Utah (links: 1924).
Diese Abdrücke (Trittsiegel) wurden jedoch vom Autor nicht als Beweis für die Koexistenz von Dinosauriern und Menschen herangezogen, da es sich um isoliert liegende Einzelabdrücke handelt, wie Kuban selbst zeigt (Abb. 11). Neben diesen einzelnen, fälschlicherweise als Metatarsus-Trittsiegel bezeichneten Abdrücken von Dinosauriern gibt es andere in menschlicher Schrittlänge liegende Pfade von Abdrücken, die manchmal solche ähneln, die in Abb. 12 links dargestellt sind (11) und manchmal sogar Einzelheiten wie menschenähnliche Zehen zeigen (8).
Literaturhinweise
1 Kuban, G. J. (1986): Elongate Dinosaur Tracks. This was a paper presented at the First International Conference on Dinosaur Tracks and Traces, held in Albuquerque, NM in 1986, and was subsequently published in the book Dinosaur Tracks and Traces, 1989, Gillette, D.D., and Lockley, M.G., eds., Cambridge University Press, p. 428-440. 2 Scheven, J. (1995): Leben, Nr. 6, Deutsches Schöpfungsmagazin 3 Zillmer, H.-J. (2002): Dinosaurier Handbuch. Fakten, Funde, Kontroversen, München, S. 95 ff. 4 ebd. S. 97 und Foto 2–4 5 Scherer, S. und Wiskin, R. (1986): „Menschliche Fußabdrücke“ in der Kreide: Ein Lehrstück für die Schöpfungsforschung, Arbeitsblatt WORT UND WISSEN, Nr. 15, Mai 1986 6 Ebd. S. 5 7 Zillmer, H.-J. (2005): Die Evolutionslüge, München 2005, S. 145 ff. vgl. Cremo/Thompson (1993): Forbidden Archaeology, Badger 1993 8 Zillmer, H.-J. (1998): Darwins Irrtum, München 1998 (Foto 72-77) bzw. Darwin´s Mistake, Enkhuizen 2003 9 Zillmer, H.-J. (2002): Dinosaurier Handbuch, München 10 Spektrum der Wissenschaft, Digest 5, S. 55 11 Zillmer, H.-J. (2001): Irrtümer der Erdgeschichte, München, 2001. 2. Aufl. 2003, Fotos 42-51